Introduction:
The histology of the cardiovascular system plays the main role in its function. The cardiovascular function consists of the heart, arteries, microvascular bed, veins and lymphatic vessels. These components have the main role to exchange oxygen, carbon dioxide, metabolites, salts, water and metabolic waste products.
The heart (Fig. 1):
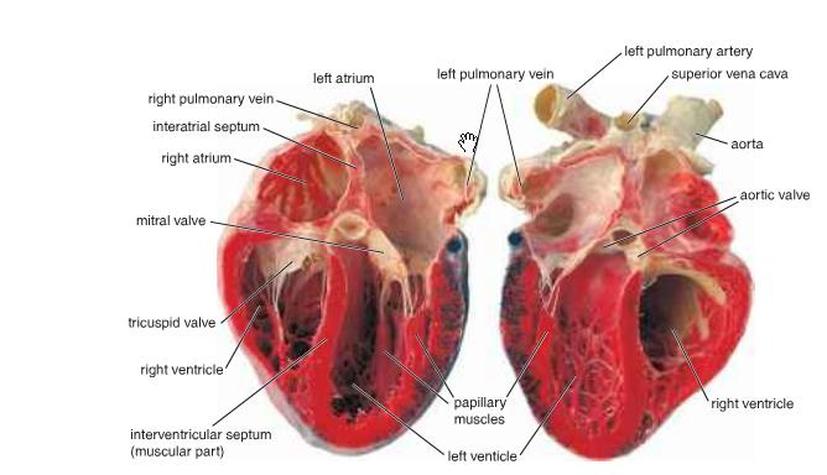
Figure 1: Heart structure ( Ross and Pawlina, 2011 ).
The heart wall consists of three layers (Fig. 2 a, b and c):
The first layer is the epicardium, which is the visceral layer of the serous pericardium. This layer covers the outer layer of the heart and consists of a single layer of mesothelial cells and connective tissue. Also, it consists of adipose tissue, which surrounds the blood vessels and nerves that supply the heart. At the entry of the great vessels, the epicardium reflects back as the parietal layer of the serous pericardium, which lines the inner surface of the pericardial sac that covers the heart surface and the roots of the great vessels. Consequently, between the visceral layer and the parietal layers of the serous pericardium is a space known as the pericardial cavity, which is filled with 15-50 ml of serous pericardial fluid.
The second layer is the myocardium, which consists of the cardiac muscle. Because the atria receive blood from the large veins and then delivers it to the ventricles and this process requires low pressure, the atrial wall is thinner than the walls of the ventricles.
The first layer is the epicardium, which is the visceral layer of the serous pericardium. This layer covers the outer layer of the heart and consists of a single layer of mesothelial cells and connective tissue. Also, it consists of adipose tissue, which surrounds the blood vessels and nerves that supply the heart. At the entry of the great vessels, the epicardium reflects back as the parietal layer of the serous pericardium, which lines the inner surface of the pericardial sac that covers the heart surface and the roots of the great vessels. Consequently, between the visceral layer and the parietal layers of the serous pericardium is a space known as the pericardial cavity, which is filled with 15-50 ml of serous pericardial fluid.
The second layer is the myocardium, which consists of the cardiac muscle. Because the atria receive blood from the large veins and then delivers it to the ventricles and this process requires low pressure, the atrial wall is thinner than the walls of the ventricles.
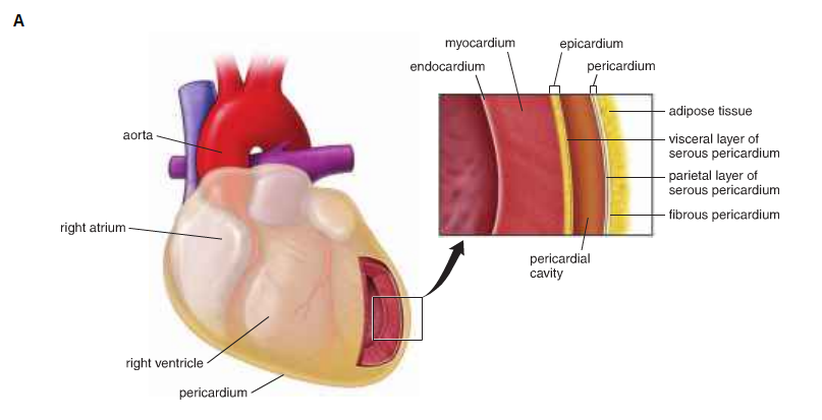
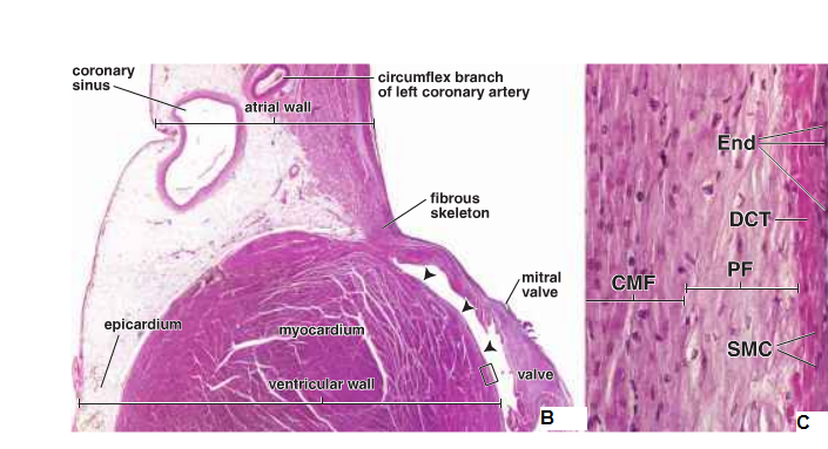
Figure 2: The layers of the heart wall. End: endothelium, DCT: dense connective tissue, PF: Purkinje fibres, CMF: cardiac muscle fibres and SMC: smooth muscle cells ( Ross and Pawlina, 2011 ).
The histological structure of the cardiac muscle (Fig. 3):
The first characteristic feature of cardiac muscle is the central location of the nucleus, which helps to differentiate the cardiac muscle from the multinucleated skeletal muscle. In addition, in the atria of the heart there are atrial granules, which contain two polypeptide hormones, atrial natriuretic factor (ANF) and brain natriuretic factor (BNF).
Additionally, in the juxtanuclear region there are large mitochondria that are densely packed between the myofibrils. These mitochondria release and recapture energy. Also, glycogen granules are concentrate between the myofibrils: these granules help to store energy in the cardiac muscle.
The third feature is the intercalated disc, which consists of transverse and the lateral components. These components have specialised cell-to-cell junctions between adjoining cardiac muscle cells:
1- Fascia adherens: This is the main junction of the transverse component of the intercalated disc and it can be stained by routine H&E. It holds the cardiac muscle cells together at their ends to form the cardiac muscle fibres.
2- Maculae adherens: These bind the individual muscle cells to one another. This prevents the cells from pulling apart under the strain of regular repetitive contractions.
3- Gap junction: This is a major structural element of the lateral component of the intercalated disc. It provides ionic continuity between adjacent cardiac muscle cells.
The third layer of the heart wall is the endocardium, which consists of an inner layer of endothelium and subendothelial connective tissue, a middle layer of connective tissue and smooth muscle cells and a deeper layer of connective tissue, which is known as the subendocardial layer and is continuous with the connective tissue of the myocardium. The conducting system is located in the subendocardial layer of the endocardium.
The endocardium lines each surface of the interventricular septum, which contains cardiac muscle, except for the membranous portion. The interatrial septum is thinner than the interventricular septum, except in some areas which contain fibrous tissue.
Additionally, in the juxtanuclear region there are large mitochondria that are densely packed between the myofibrils. These mitochondria release and recapture energy. Also, glycogen granules are concentrate between the myofibrils: these granules help to store energy in the cardiac muscle.
The third feature is the intercalated disc, which consists of transverse and the lateral components. These components have specialised cell-to-cell junctions between adjoining cardiac muscle cells:
1- Fascia adherens: This is the main junction of the transverse component of the intercalated disc and it can be stained by routine H&E. It holds the cardiac muscle cells together at their ends to form the cardiac muscle fibres.
2- Maculae adherens: These bind the individual muscle cells to one another. This prevents the cells from pulling apart under the strain of regular repetitive contractions.
3- Gap junction: This is a major structural element of the lateral component of the intercalated disc. It provides ionic continuity between adjacent cardiac muscle cells.
The third layer of the heart wall is the endocardium, which consists of an inner layer of endothelium and subendothelial connective tissue, a middle layer of connective tissue and smooth muscle cells and a deeper layer of connective tissue, which is known as the subendocardial layer and is continuous with the connective tissue of the myocardium. The conducting system is located in the subendocardial layer of the endocardium.
The endocardium lines each surface of the interventricular septum, which contains cardiac muscle, except for the membranous portion. The interatrial septum is thinner than the interventricular septum, except in some areas which contain fibrous tissue.
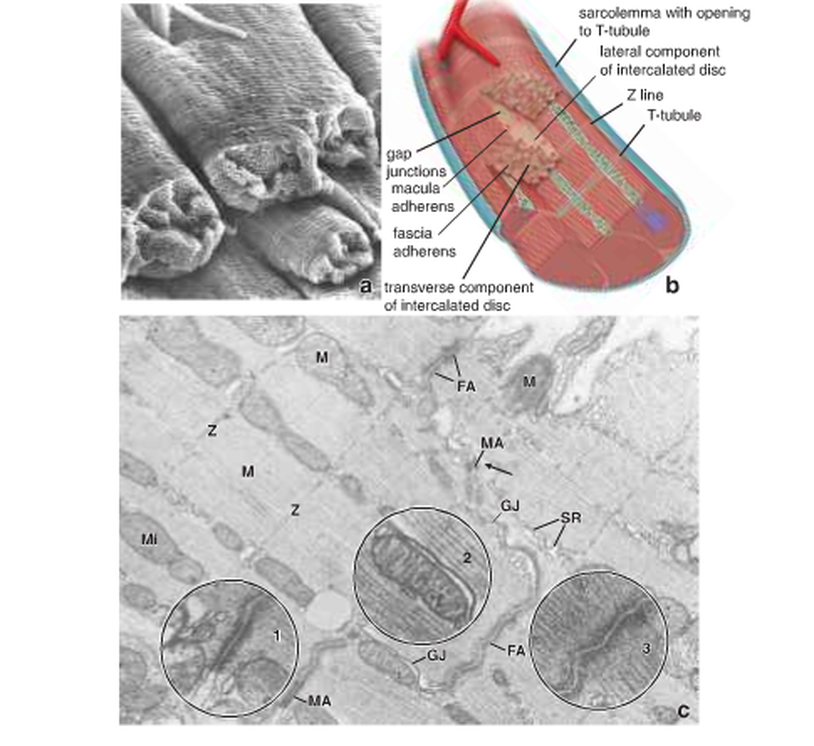
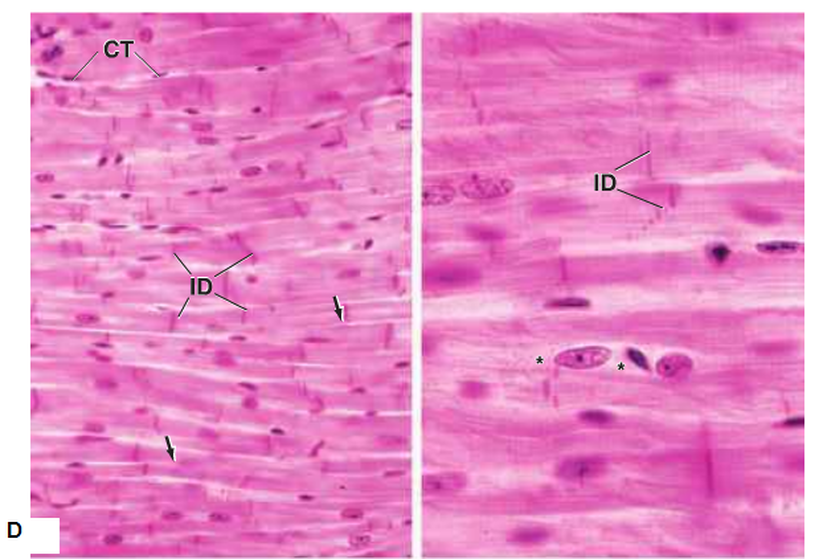
Figure 3: The histological structure of the cardiac muscle. CT: connective tissue, ID: intercalated discs ( Ross and Pawlina, 2011 )
.
.
Heart valves (Fig. 4):
The heart valves are composed of dense irregular connective tissue, which forms fibrous rings to surround the orifices. Each valve has three layers:
1- Fibrosa, which forms the core of the valve. This layer has fibrous extensions from the dense irregular connective tissue of the skeletal rings of the heart.
2- Spongiosa, which is loosely arranged collagen and elastic fibres infiltrated with large numbers of proteoglycans. This layer is located on the atrial or blood vessel side of each valve. In the aortic and pulmonary vessels the valves are called arterialis; however, in the AV (tricuspid and mitral) vessels, the valves are called auricularis.
The ventricularis, which is adjacent to the ventricular or atrial surface of each valve. The endothelium covers this layer. Also, this layer contains dense connective tissue with many layers of elastic fibres.
1- Fibrosa, which forms the core of the valve. This layer has fibrous extensions from the dense irregular connective tissue of the skeletal rings of the heart.
2- Spongiosa, which is loosely arranged collagen and elastic fibres infiltrated with large numbers of proteoglycans. This layer is located on the atrial or blood vessel side of each valve. In the aortic and pulmonary vessels the valves are called arterialis; however, in the AV (tricuspid and mitral) vessels, the valves are called auricularis.
The ventricularis, which is adjacent to the ventricular or atrial surface of each valve. The endothelium covers this layer. Also, this layer contains dense connective tissue with many layers of elastic fibres.
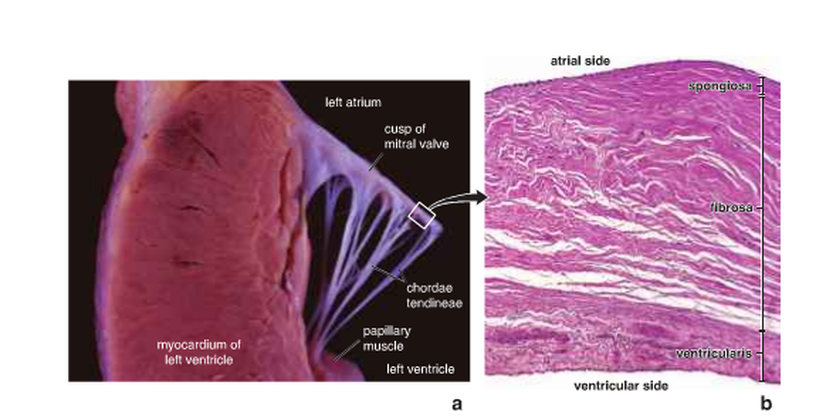
Figure 4: a) a cuspid valve; b) tissue layers within heart valves (Ross and Pawlina, 2011).
Specialised cells of the conducting system(Fig. 5 a & b):
Cell nodes:
The SA-node and AV-node have specialised fusiform cells, which are smaller than cardiac muscle cells. However, they are similar with reference to myofibril and extrafibrillar sarcolemma, with the exception of the transverse tubules of the T-system. These cells differ in their arrangement because they are mutually interconnected side to side because of their fusiform shape and they form the cord like clusters of cells. Although intercalated disks do not occur, all three of the junctional specialisations of sarcolemma are distributed along the surfaces in contact. Furthermore, a thin basal lamina and a thin layer of connective tissue fibrils invest the cord-like clusters of specialised cells and the specialised muscle cells of the nodal tissue in contact with the atrial muscle cells.
Cells of AV bundle and branches:
The major part of the AV bundle is made of similar cells, as found in the nodes. However, in the distal parts of the bundle and throughout the interventricular branches change from fusiform to elongated cylindrical shapes and they meet end to end. These cells known are as Purkinje fibres.
The SA-node and AV-node have specialised fusiform cells, which are smaller than cardiac muscle cells. However, they are similar with reference to myofibril and extrafibrillar sarcolemma, with the exception of the transverse tubules of the T-system. These cells differ in their arrangement because they are mutually interconnected side to side because of their fusiform shape and they form the cord like clusters of cells. Although intercalated disks do not occur, all three of the junctional specialisations of sarcolemma are distributed along the surfaces in contact. Furthermore, a thin basal lamina and a thin layer of connective tissue fibrils invest the cord-like clusters of specialised cells and the specialised muscle cells of the nodal tissue in contact with the atrial muscle cells.
Cells of AV bundle and branches:
The major part of the AV bundle is made of similar cells, as found in the nodes. However, in the distal parts of the bundle and throughout the interventricular branches change from fusiform to elongated cylindrical shapes and they meet end to end. These cells known are as Purkinje fibres.
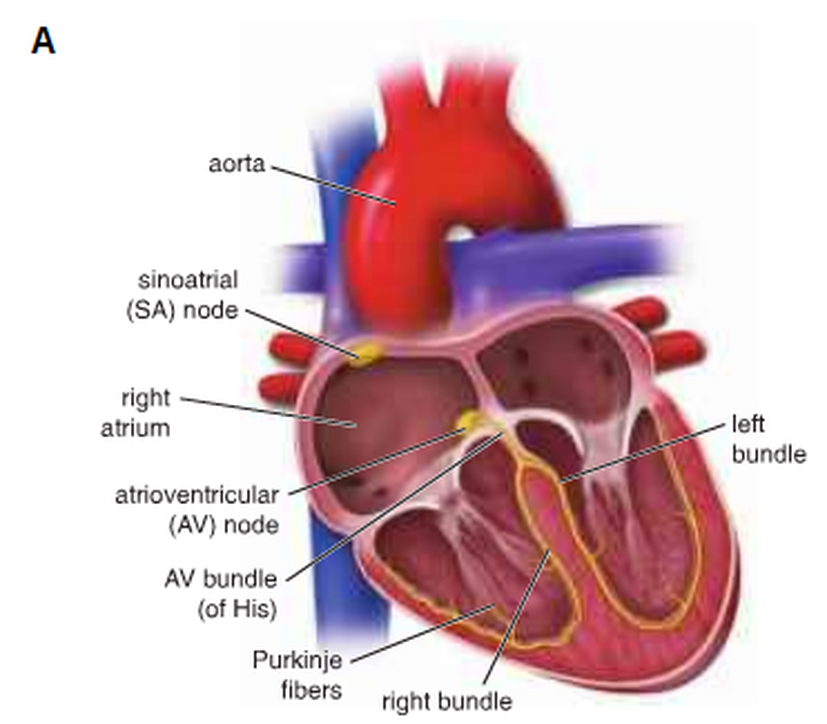
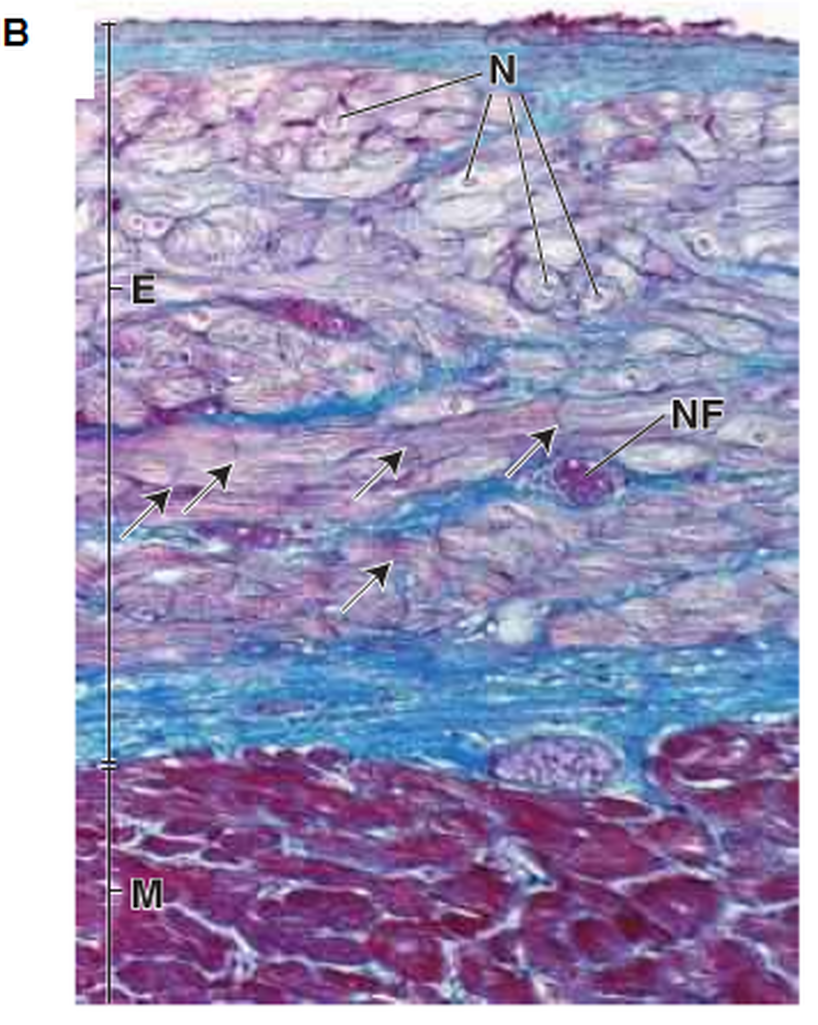
Figure 5: The conducting system. N: nuclei, E: endocardium, NF: Purkinje fibres course nerves and M: myocardium (Ross and Pawlina, 2011).
Lymphatic vessels (Fig. 6):
The lymphatic vessels are filled with lymph and run beside the blood vessels. The smallest lymphatic vessels are known as lymphatic capillaries and are numerous under the epithelium of the skin and mucous membrane. These capillaries form the larger vessels called lymphatic vessels, which unite to form two main channels that empty into the large veins in the base of the neck on the left side. This largest lymphatic vessel is known as the thoracic duct and the other important channel is known as the right lymphatic trunk. The lymphatic capillaries have endothelium, which lacks a continuous basal lamina; between the incomplete basal lamina and the perivascular collagen extend the anchoring filaments. The thickness of the lymphatic vessels increases because of the connective tissue and bundles of smooth muscle. The lymphatic vessels also have valves to prevent backflow of the lymph, which moves due to the compression of the lymphatic vessels by adjacent skeletal muscles.
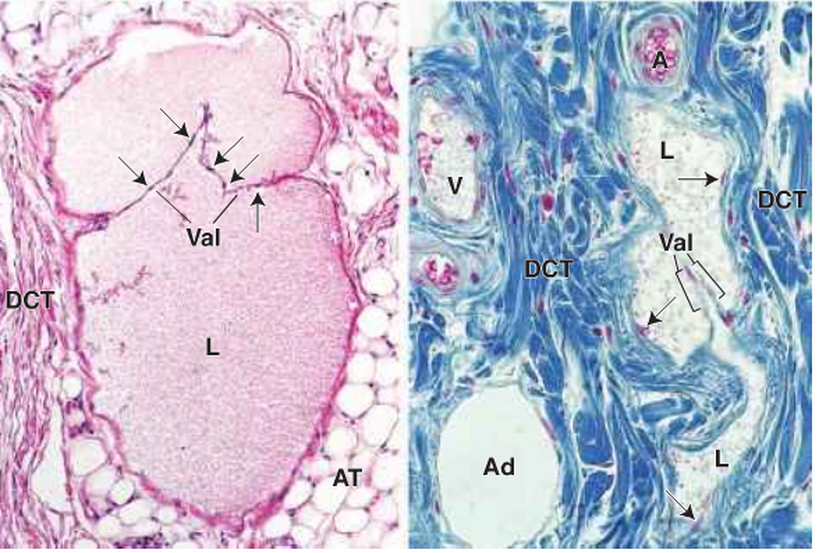
Figure 6: Lymphatic vessels. Val: valve, DCT: dense irregular connective tissue, AT: adipose tissue, Ad: adipocyte, A: arteriole, L: lymph material and V: venule (Ross and Pawlina, 2011).
Arteries:
The arteries are divided into three types:
Large or elastic arteries (Fig. 7, 8 & 10):
These have multiple sheets of elastic lamella in their walls. Firstly, the tunica intima of elastic arteries have endothelium with basal lamina. The cells are flat and elongated in shape and are joined by a tight junction (zonulae occuludentes) and gap junction during formation of the epithelial sheet. Also, their cytoplasm contain weibel-palade bodies. The endothelial organelles contain von Willebrand factor and P-selectin. During endothelial injury, the Von Willebrand factor, which is a synthesised glycoprotein, is secreted into the blood and binds to the coagulating factor VIII. This plays the major role in the adhesion of platelets. In addition, the tunica intima consists of the subendothelial layer, which consists of connective tissue with both collagen and elastic fibres. Finally, the tunica intima consists of an internal elastic membrane.
Secondly, the tunica media is the thickest of the three layers in elastic arteries. It consists of elastin, smooth muscle cells, collagen fibres and ground substance, which are synthesised and secreted by smooth muscle cells. Fibroblasts are not present in the tunica media. Thirdly, the tunica adventitia consists of a thin connective tissue layer. For example, collagen fibres and elastic fibres form a loose network of elastic fibres. Also, the tunica adventitia consists of fibroblasts, macrophages, vasa vasorum and nervi vascularis. The last two layers have similar arterial and veins that branch to provide nutrients and remove waste products from the outer portion of the arterial wall.
Large or elastic arteries (Fig. 7, 8 & 10):
These have multiple sheets of elastic lamella in their walls. Firstly, the tunica intima of elastic arteries have endothelium with basal lamina. The cells are flat and elongated in shape and are joined by a tight junction (zonulae occuludentes) and gap junction during formation of the epithelial sheet. Also, their cytoplasm contain weibel-palade bodies. The endothelial organelles contain von Willebrand factor and P-selectin. During endothelial injury, the Von Willebrand factor, which is a synthesised glycoprotein, is secreted into the blood and binds to the coagulating factor VIII. This plays the major role in the adhesion of platelets. In addition, the tunica intima consists of the subendothelial layer, which consists of connective tissue with both collagen and elastic fibres. Finally, the tunica intima consists of an internal elastic membrane.
Secondly, the tunica media is the thickest of the three layers in elastic arteries. It consists of elastin, smooth muscle cells, collagen fibres and ground substance, which are synthesised and secreted by smooth muscle cells. Fibroblasts are not present in the tunica media. Thirdly, the tunica adventitia consists of a thin connective tissue layer. For example, collagen fibres and elastic fibres form a loose network of elastic fibres. Also, the tunica adventitia consists of fibroblasts, macrophages, vasa vasorum and nervi vascularis. The last two layers have similar arterial and veins that branch to provide nutrients and remove waste products from the outer portion of the arterial wall.
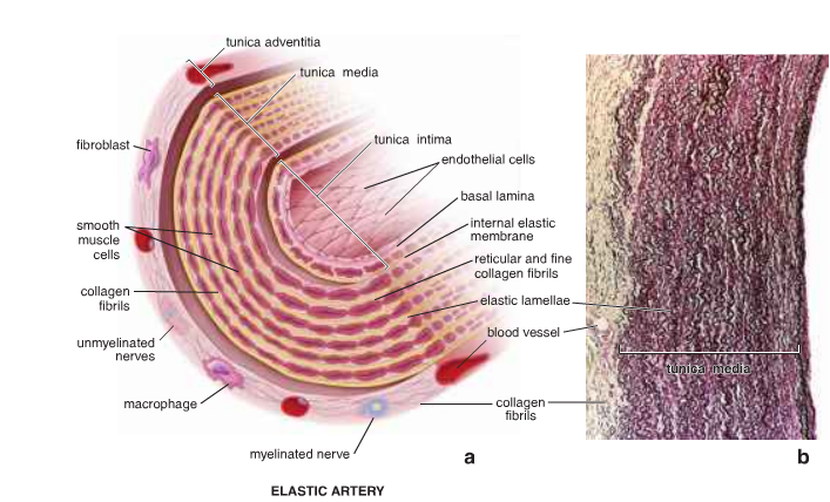
Figure 7: The structure of the large and elastic artery( Ross and Pawlina, 2011 ).
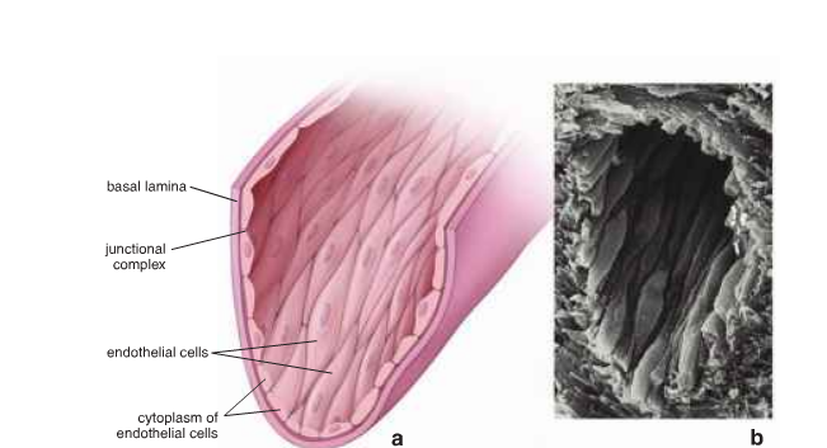
Figure 8: The morphology of the endothelium( Ross and Pawlina, 2011 ).
Medium arteries or muscular arteries (Fig. 9 & 10):
Firstly, the tunica intima is thinner than in elastic arteries. These arteries consist of an endothelial lining with a basal lamina, a subendothelial layer of connective tissue and an internal elastic membrane, which contacts the basal lamina of the endothelium. Secondly, the tunica media is composed of smooth muscle and less elastin than the elastic arteries. Thirdly, the tunica adventitia is thick and separated from the tunica media by a recognisable external elastic membrane.
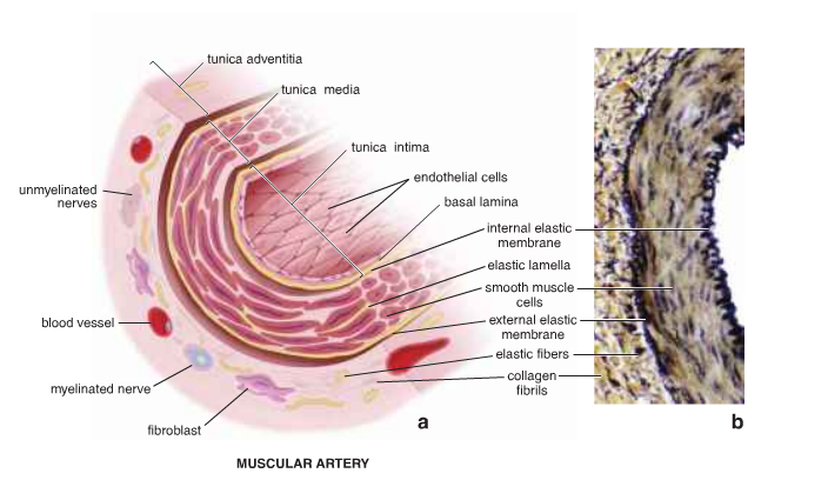
Figure 9: The structure of the muscular artery ( Ross and Pawlina, 2011 ).
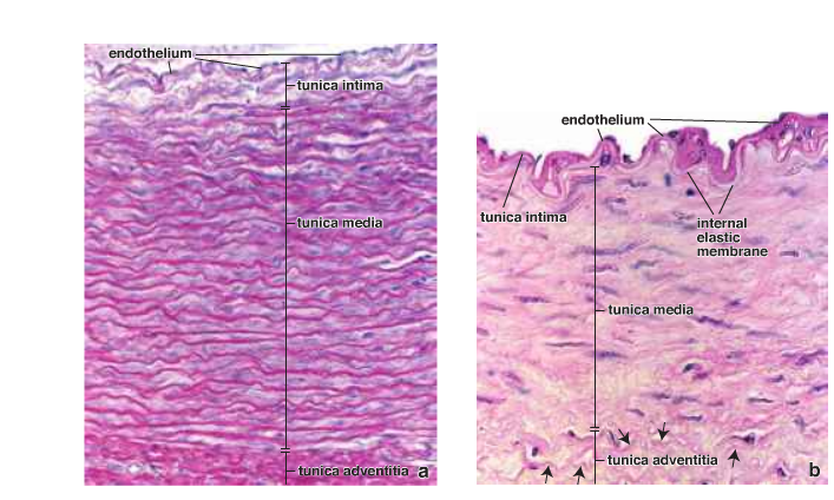
Figure 10: The wall of the elastic artery (a) and muscular artery (b) ( Ross and Pawlina, 2011 ).
Small arteries and arterioles (Fig. 11):
There are slight differences between small arteries and arterioles in their layers. Firstly, the tunica intima in small arteries has an internal elastic membrane; however, the arterioles may or may not have this layer. The endothelium is similar to the other arteries. Secondly, in the tunica media the arterioles have only one or two layers of smooth muscle; however, the small arteries may have eight layers of smooth muscle. Finally, the tunica adventitia is a thin and ill-defined sheath of connective tissue, which blends with the connective tissue.
There are slight differences between small arteries and arterioles in their layers. Firstly, the tunica intima in small arteries has an internal elastic membrane; however, the arterioles may or may not have this layer. The endothelium is similar to the other arteries. Secondly, in the tunica media the arterioles have only one or two layers of smooth muscle; however, the small arteries may have eight layers of smooth muscle. Finally, the tunica adventitia is a thin and ill-defined sheath of connective tissue, which blends with the connective tissue.
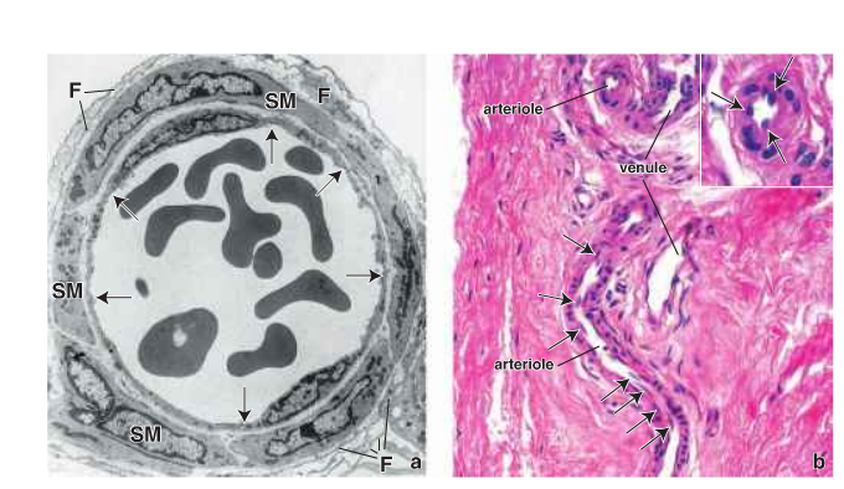
Figure 11: The structure of the arterioles. F: fibroblast and SM: smooth muscle ( Ross and Pawlina, 2011 ).
Capillaries (Fig. 12):
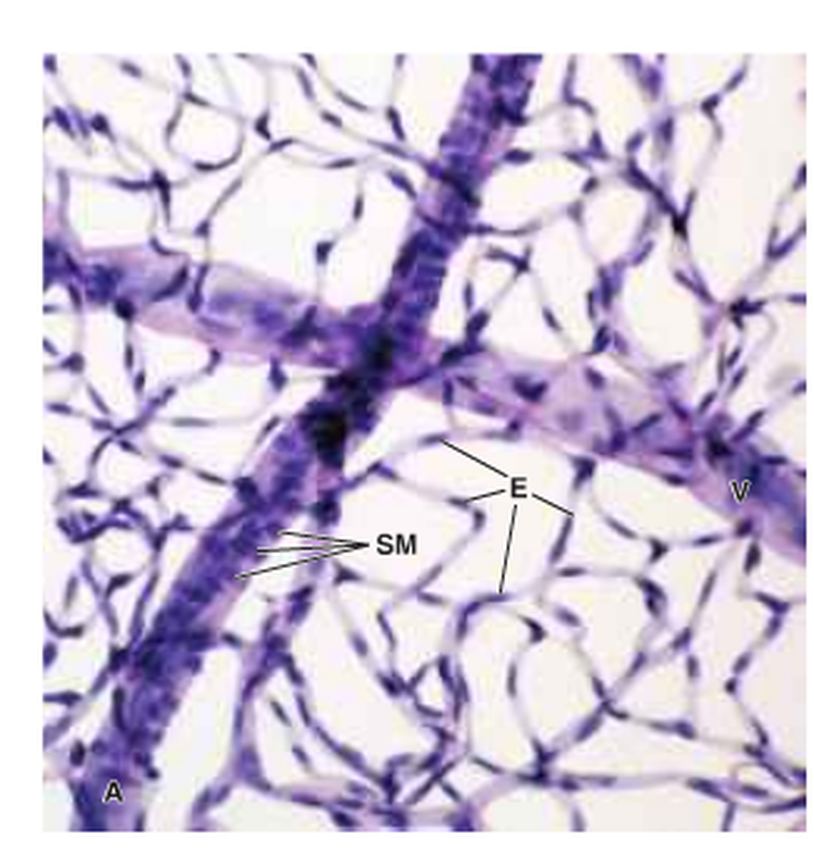
Figure 12: The structure of the capillary network. SM: smooth muscle cells, E: nuclei of endothelial cells and V: venule ( Ross and Pawlina, 2011 ).
Their diameter is smaller than the erythrocytes. They are divided into three types:
The first type consists of continuous capillaries (Fig. 13), which are found in muscle, lungs and the central nervous system (CNS). In cross section these capillaries appear as two plasma membranes enclosing a ribbon of the cytoplasm with occluding junctions. Also, pericytes/rouget cells are associated with the endothelium. These cells surround the capillaries with branching cytoplasmic processes and are enclosed by a basal lamina, which is continuous with the endothelium.
The first type consists of continuous capillaries (Fig. 13), which are found in muscle, lungs and the central nervous system (CNS). In cross section these capillaries appear as two plasma membranes enclosing a ribbon of the cytoplasm with occluding junctions. Also, pericytes/rouget cells are associated with the endothelium. These cells surround the capillaries with branching cytoplasmic processes and are enclosed by a basal lamina, which is continuous with the endothelium.
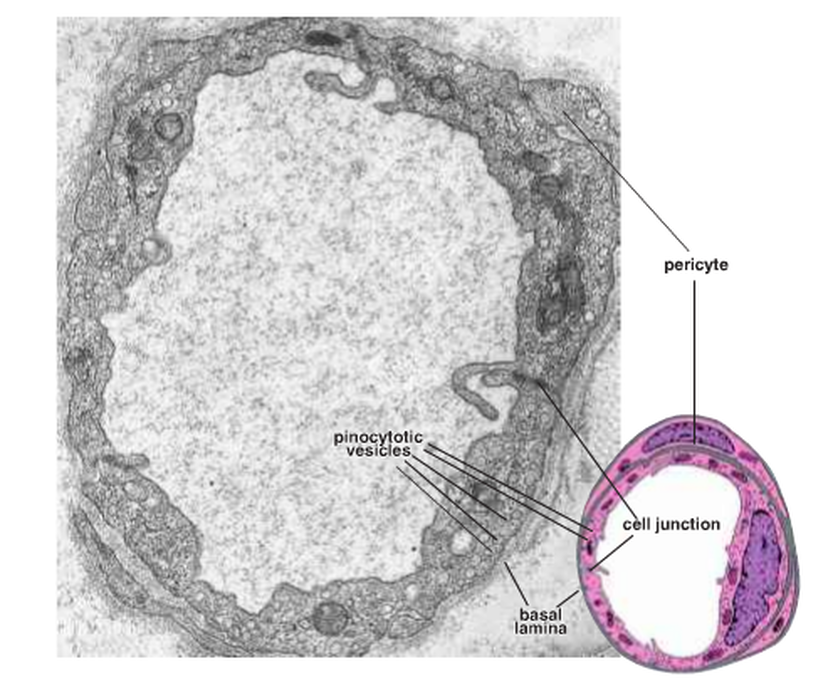
Figure 13: The continuous capillaries ( Ross and Pawlina, 2011 ).
The second type is the fenestrated capillaries (Fig. 14), which are found in the endocrine glands and the sites of metabolic and fluid absorption; for example, the gallbladder, kidney and intestinal tract. These capillaries have fenestration channels across the capillary wall and pinocytotic vesicles.
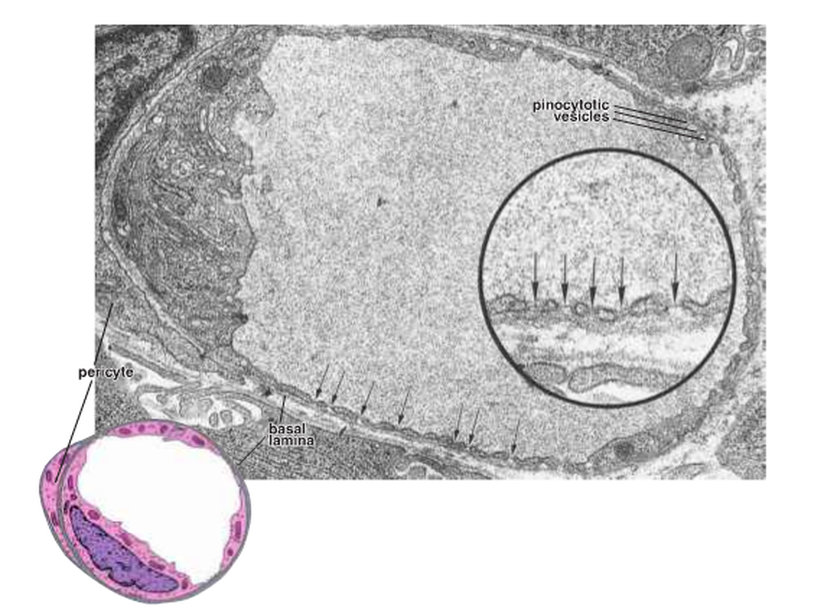
Figure 14: The fenestrated capillaries ( Ross and Pawlina, 2011 ).
The third type consists of discontinuous capillaries, which are also known as sinusoidal capillaries or sinusoids. These types are found in the liver, spleen and bone marrow. Their diameter is larger than other types and they have irregular shape. They vary from organ to organ; for instance, in the liver the Kupffer cells and vitamin A-storing Ito cells are associated with the endothelial cells. In the spleen, the endothelial cells are spindle- shape with gaps between the neighbouring cells: the basal lamina underlying the endothelium may be partially or completely absent.
Arteriovenous shunts (Fig. 15):
There are routes called arteriovenous (AV) anastomoses or AV shunts between arteries and veins to allow the blood to bypass capillaries. These are found in the skin, fingertips, nose, lips and erectile tissue of the penis and clitoris. The arteriole of AV shunts is coiled and has a thick smooth muscle layer near to the connective tissue capsule and many nerves.
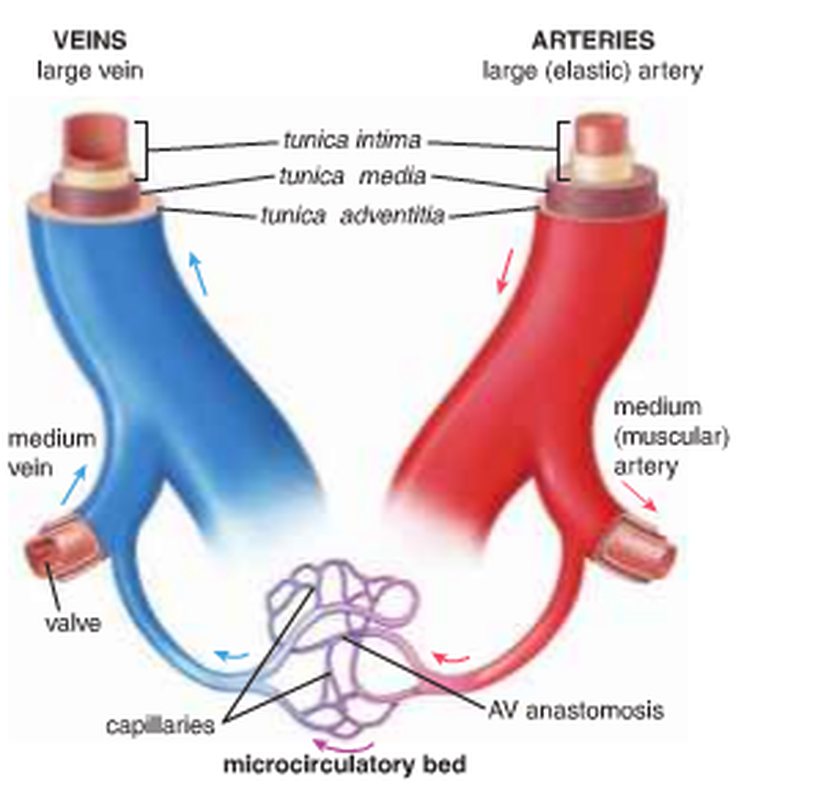
Figure 15: AV shunts ( Ross and Pawlina, 2011 ).
Veins:
Veins are classified in three types:
Venules and small veins:
This type has two subtypes. Firstly, postcapillary venules have an endothelial lining with a basal lamina and pericytes, which form an umbrella where they connect with endothelial cells. The endothelial cells and pericytes synthesise and share the basal lamina. In lymph nodes, the postcapillary venules have a cuboidal appearance and ovoid nuclei; as a result they are known as high endothelial venules (HEVs). Secondly, there are muscular venules, which are located in the distal to the postcapillary venules. This type has one or two layers of smooth muscle, which form the tunica media; however, the postcapillary venules have no true tunica media. Also, there are no pericytes in muscular venules.
Medium veins (Fig. 16):
The deepest veins that are associated with arteries are medium veins. This type has three tunics in the venous wall:
1- Tunica intima, which consists of endothelium with its basal lamina, a thin subendothelial layer with smooth muscle cells and may consist of a thin internal elastic membrane.
2- Tunica media, a thinner layer than in medium-sized arteries. It has several layers of smooth muscle with interspersed collagen and elastic fibres.
3- Tunica adventitia, which is thicker than the tunica media. It has collagen fibres and networks of elastic fibres.
Venules and small veins:
This type has two subtypes. Firstly, postcapillary venules have an endothelial lining with a basal lamina and pericytes, which form an umbrella where they connect with endothelial cells. The endothelial cells and pericytes synthesise and share the basal lamina. In lymph nodes, the postcapillary venules have a cuboidal appearance and ovoid nuclei; as a result they are known as high endothelial venules (HEVs). Secondly, there are muscular venules, which are located in the distal to the postcapillary venules. This type has one or two layers of smooth muscle, which form the tunica media; however, the postcapillary venules have no true tunica media. Also, there are no pericytes in muscular venules.
Medium veins (Fig. 16):
The deepest veins that are associated with arteries are medium veins. This type has three tunics in the venous wall:
1- Tunica intima, which consists of endothelium with its basal lamina, a thin subendothelial layer with smooth muscle cells and may consist of a thin internal elastic membrane.
2- Tunica media, a thinner layer than in medium-sized arteries. It has several layers of smooth muscle with interspersed collagen and elastic fibres.
3- Tunica adventitia, which is thicker than the tunica media. It has collagen fibres and networks of elastic fibres.
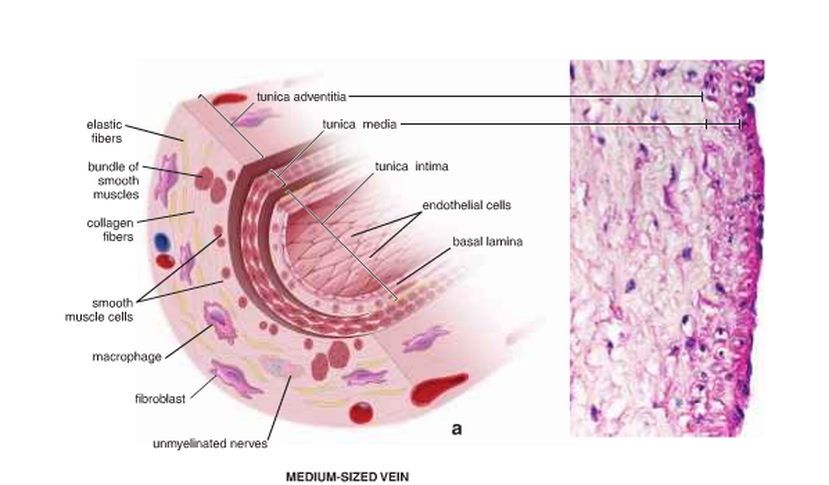
Figure 16: The structure of the medium veins( Ross and Pawlina, 2011 ).
Large veins (Fig. 17):
These vessels have a diameter larger than 10 mm, such as the venae cavae, pulmonary vein, innominates, azygos, renal, adrenal, splenic and superior mesenteric veins. Their walls have three layers. Firstly, the tunica intima consists of an endothelial lining with basal lamina, a small amount of subendothelial connective tissue and smooth muscle cells, which are close to the intimal endothelium. These muscle cells may belong to the tunica intima or the tunica media. Secondly, the thin layer is the tunica media, which contains smooth muscle, collagen fibres and some fibroblasts. Finally, the thickest layer of the vessel wall is the tunica adventitia, which consists of collagen, elastic fibres, fibroblasts and longitudinally disposed smooth muscle cells.
These vessels have a diameter larger than 10 mm, such as the venae cavae, pulmonary vein, innominates, azygos, renal, adrenal, splenic and superior mesenteric veins. Their walls have three layers. Firstly, the tunica intima consists of an endothelial lining with basal lamina, a small amount of subendothelial connective tissue and smooth muscle cells, which are close to the intimal endothelium. These muscle cells may belong to the tunica intima or the tunica media. Secondly, the thin layer is the tunica media, which contains smooth muscle, collagen fibres and some fibroblasts. Finally, the thickest layer of the vessel wall is the tunica adventitia, which consists of collagen, elastic fibres, fibroblasts and longitudinally disposed smooth muscle cells.
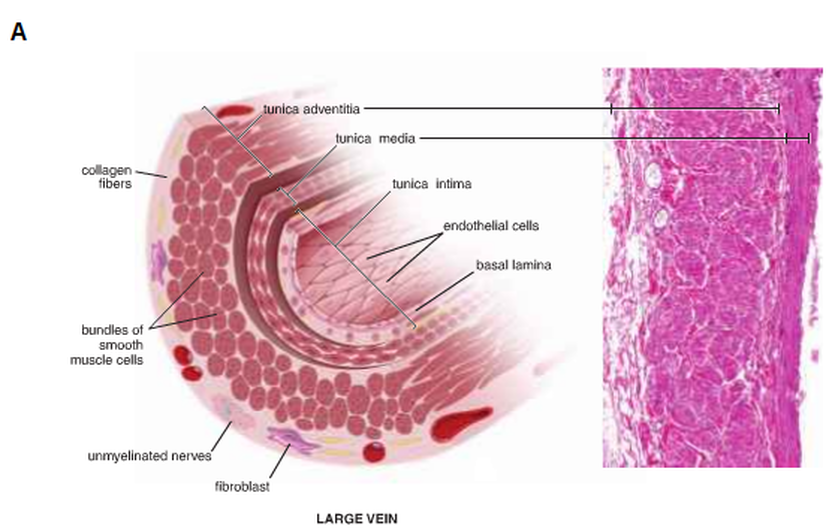
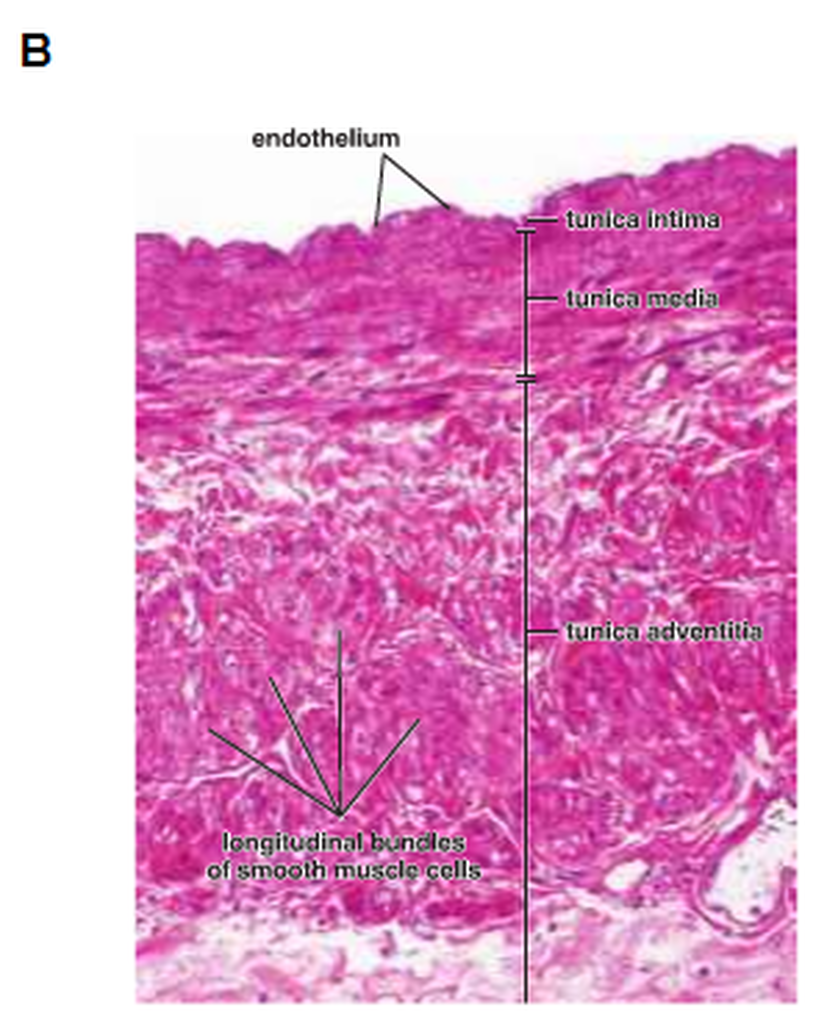
Figure 17: The structure of the large vein ( Ross and Pawlina, 2011 ).
Copyright © 2011 by Reem Bu Saeed.
Last updated: 30-07-2011.
Last updated: 30-07-2011.
